
Миома матки (лейомиома, ЛММ) – доброкачественная гормонозависимая опухоль, развивающаяся из мышечных и соединительнотканных элементов, которая относится к группе мезенхимальных опухолей [1]. Узлы опухоли чаще развиваются в местах наиболее сложных переплетений мышечных волокон – по средней линии матки, вблизи трубных углов, по бокам шейки матки. Одновременно с ростом узлов миомы увеличивается масса окружающего миометрия, но темп роста опухоли существенно опережает таковой разрастания миометрия. Миоматозные узлы не имеют капсулы и окружены гипертрофированным мышечным слоем матки. Предполагается, что одним из основных факторов, вызывающих развитие локальной гипертрофии миометрия, является гипоксия как результат микроциркуляторных нарушений, что обусловливает компенсаторную перестройку гладкомышечных клеток с последующими дистрофическими изменениями и секрецией ими компонентов экстрацеллюлярного матрикса. В узлах, локализующихся в среднем и внутреннем слоях миометрия, соотношение паренхимы и стромы в среднем составляет 2 : 1, подбрюшинных узлах – 1 : 3. Опухолевый узел повторяет в своем развитии паренхиматозно-стромальные особенности того слоя миометрия, из которого он развивается [14].
Кроме разделения опухолей по тканевому признаку выделяют простые и пролиферирующие опухоли; последние встречаются почти у каждой четвертой женщины с миомой матки. Мышечные клетки в пролиферирующих миомах более многочисленны по сравнению с миомами без признаков пролиферации. В пролиферирующих миомах нередко возрастает число тучных, плазматических и лимфоидных клеток.
Причина формирования ЛММ остается неизвестной. В настоящее время доказанными этиопатогенетическими факторами развития ЛММ являются половые стероиды, тканевые гормоны роста и генетическая предрасположенность (нерандомизированные хромосомные альтерации). Иерархия перечисленных факторов активно обсуждается [4].
Цитогенетические исследования ЛММ
Поиск причины возникновения ЛММ закономерно привел к попыткам выявления возможных генетических аномалий. В пользу наследственной природы заболевания говорят эпидемиологические данные о высокой частоте ЛММ у представительниц афроамериканской расы и совпадении частотных данных у близких родственниц. Микроскопический анализ позволил выявить опухолеспецифические кариотипические изменения в ткани лишь 40–60 % миоматозных узлов (Rein M.S., 1991). При этом наличие нормального кариотипа не исключает субмикроскопических генетических аберраций. Тем не менее пусковой механизм генетических изменений при инициации неогенеза окончательно не определен. Наиболее часто аберрации затрагивают хромосомы 6, 7, 12 и 14. Как и предполагалось, изменения наблюдаются в области локализации генов, ответственных за процессы деления, дифференцировки и апоптоза. В настоящее время установлена клоновая и мозаичная природа ЛММ, выявлена корреляция гено- и фенотипических особенностей, определены цитогенетические подгруппы и идентифицированы отдельные гены, вовлеченные в канцерогенез [35].
Миометриальные гладкомышечные клетки с приобретенными мутациями проявляют преимущественный рост перед интактными клетками, активно пролиферируют за счет повышенной чувствительности к вне- и внутриклеточным стимуляторам роста. Догма о том, что миома вырастает в результате экспансии одной клеткипредшественницы с генетической поломкой была поколеблена Mashal R.D.(1994) и Baschinsky D.Y. (2000), доказавшими, что хромосомная поломка может развиваться вторично при моноклональной экспансии предсуществующей ЛММ [12, 37].
Исследователи постоянно предлагают новые гены-кандидаты, которые, по их мнению, являются первопричиной заболевания. В данном ряду последовательно рассмотрены гены, кодирующие фумаратгидратазу (Gross K.L. и соавт., 2004), катехол-О-метилтрансферазу, группы высокомобильных генов белков HMGIC и HMGIY [50], туберина, рецепторов глюкокортикоидов, кофакторов стероид-зависимой транскрипции [55].
Тем не менее ни один из выявленных генов, экспрессия которых изменена в ЛММ, не может быть единственным ответственным за инициацию/развитие ЛММ. Это связано с гетерогенной природой ЛММ, когда прослеживается связь со специфическими, но разными генами. Другими словами, ЛММ представлена несколькими отдельными цитогенетическими субгруппами, что подтверждает мнение о множественных генетических путях, ведущих к канцерогенезу. При этом частота встречающихся аберраций говорит о том, что вряд ли это первичный патогенетический эффект.
Интересны недавние исследования микро-РНК в миоматозной ткани [9]. Микро-РНК отвечают за стабильность экспрессии генов и представляют собой семейство малых некодирующих белков РНК. Они присутствуют во всех хромосомах, за исключением Y. Микро-РНК комплементарно связывается
с 3-концом гена-мишени и регулирует его транскрипцию, а также дальнейшую трансляцию продукта. У человека идентифицировано более 1500 микро-РНК. При этом более трети генов человека содержит участки связывания для микро-РНК. Хотя биологическое значение для большинства микро-РНК остается неизвестным, последние исследования доказывают их участие в регуляции клеточного цикла, апоптоза и дифференцировки. В целом можно сказать, что при миоме наблюдается аберрантная экспрессия микро-РНК.
с 3-концом гена-мишени и регулирует его транскрипцию, а также дальнейшую трансляцию продукта. У человека идентифицировано более 1500 микро-РНК. При этом более трети генов человека содержит участки связывания для микро-РНК. Хотя биологическое значение для большинства микро-РНК остается неизвестным, последние исследования доказывают их участие в регуляции клеточного цикла, апоптоза и дифференцировки. В целом можно сказать, что при миоме наблюдается аберрантная экспрессия микро-РНК.
Pan Q. (2010) проанализировал в ткани миомы матки уровень 27 микроРНК, о которых известно, что они задействованы в регуляции клеточного цикла, дифференцировке, подвижности клеток, апоптозе и обмене компонентов внеклеточного матрикса. Четыре из этих микро-РНК предположительно регулируют экспрессию генов рецепторов эстрадиола и прогестерона. Надо сразу отметить, что присвоение отдельной микро-РНК определенной узкой функции невозможно, т. к. одна микро-РНК потенциально может регулировать экспрессию сотни генов, и наоборот – каждый ген может служить потенциальной мишенью для нескольких разных микро-РНК. Одна из микро-РНК, экспрессия которой изменяется при опухолевой трансформации миометрия, MiR-21, контролирует экспрессию рецептора трансформирующего фактора роста-β 2-го типа (ключевого профибротического цитокина, который вызывает гипертрофию клеток, нарушает обмен компонентов межклеточного матрикса и влияет на ангиогенез). Наиболее интересно взаимодействие экспрессии микр-РНК и рецепторов половых стероидов. Половые стероиды регулируют экспрессию микро-РНК, т. к. последняя зависит от фазы менструального цикла. Эстрадиол и медроксипрогестерона ацетат (МПА) по-разному регулируют экспрессию микро-РНК в разных типах клеток. В частности, эстрадиол ингибирует, а МПА стимулирует экспрессию MiR-21 и MiR-26a в гистологически неизмененной ткани миометрия (ГНТМ) и ткани ЛММ соответственно. Таким образом, выявленная аберрантная экспрессия микро-РНК может приводить к репрограммированию экспрессии специфических генов и трансформации миофибробластных клеток в ЛММ.
Половые стероиды в патогенезе миомы матки
Гормоны плазмы крови
Многочисленные литературные данные позволяют с уверенностью утверждать, что наиболее важным фактором, контролирующим процессы гипертрофии и гиперплазии клеток миомы матки, являются половые гормоны. В пользу участия половых стероидов говорит тот факт, что ЛММ определяется преимущественно в репродуктивном возрасте и уменьшается в размерах в менопаузальном периоде. Факторы, увеличивающие экспозицию эстрогенов в организме женщины, такие как ожирение и раннее менархе, увеличивают частоту выявления ЛММ. Тотальное снижение уровня циркулирующих половых стероидов (введение агонистов гонадолиберинов) тормозит рост узлов, в то время как после отмены препарата у пациенток репродуктивного возраста в результате восстановления овариальной функции указанный рост возобновляется, часто в опережающем темпе.
Развитие гиперпластических процессов в миометрии многие авторы связывают с гиперэстрогенией(Вихляева Е.М., 2000), другие исследователи считают, что имеют значение не столько абсолютные данные концентрации эстрогенов, сколько изменение соотношения эстрогенов и прогестинов, т. е. относительная гиперэстрогенемия [15]. С другой стороны, во многих исследованиях показано, что уровни циркулирующих половых гормонов в плазме крови при наличии ЛММ находятся в пределах референсных значений и не влияют на размеры узла [19]. Аналогичные данные получены и нами [3]. Савицкий Г.А. и соавт. (2000) выяснили, что содержание эстрадиола и прогестерона в сосудах матки выше, чем в периферической венозной крови, на основании чего ими предложена схема патогенеза миомы матки, в которой приобретение клетками миометрия новых биологических свойств напрямую связано с длительными нарушениями локального гормонального гомеостаза [5]. Таким образом, эстрогены в условиях как in vivo, так и in vitro оказывают промоторное действие на рост предсуществующей миомы.
В то же время за последние годы накоплены данные о том, что и прогестерон наряду с эстрогеном стимулирует рост миомы матки. Так, in vitro прогестерон тормозит апоптоз клеток ЛММ, стимулируя экспрессию антиапоптотического белка Bcl-2 [24]. В культуре клеток ЛММ в ответ на прогестерон повышается синтез эпидермального фактора роста (в то время как эстрадиол увеличивает экспрессию его рецепторов). Данный эффект в культуре неизмененных миометриальных клеток выражен не столь значительно, как в миомных узлах (Maruo T. и соавт., 2008). По-видимому, комбинация двух половых стероидов необходима для успешной пролиферации клеток миомы.
В литературе представлено мало исследований на животных с целью выявления роли прогестерона в пролиферации клеток ЛММ.
Тем не менее данные клинических исследований показали:
• повышение митотической активности клеток миомы, особенно у молодых женщин в лютеиновую
фазу менструального цикла (высокий прогестерон);
• у женщин, получающих мифепристон (антигестаген), происходит снижение объема миомных узлов на 49 % через 3 месяца от начала лечения при параллельном снижении уровня рецепторов прогестерона;
• прогестины реверсируют действие агонистов гонадолиберинов (аГРГ) на объем миомы.
• повышение митотической активности клеток миомы, особенно у молодых женщин в лютеиновую
фазу менструального цикла (высокий прогестерон);
• у женщин, получающих мифепристон (антигестаген), происходит снижение объема миомных узлов на 49 % через 3 месяца от начала лечения при параллельном снижении уровня рецепторов прогестерона;
• прогестины реверсируют действие агонистов гонадолиберинов (аГРГ) на объем миомы.
Последнее наблюдение наводит на мысль о приоритетной роли прогестерона в прогрессировании миомы, т. к. аГРГ вызывают отчетливую гипоэстрогенемию, которая не может быть преодолена введением прогестерона (он в отличие от тестостерона не конвертирует в эстрадиол).
Половые стероиды в ткани ЛММ
Существенные различия выявлены в уровне гормонов и их рецепторов в самой ткани ЛММ. В частности, уровень эстрадиола в ткани миомы выше, чем в гистологически неизмененной ткани миометрия. Такое различие, по-видимому, связано с местным синтезом гормона. Клетки ЛММ экспрессируют два фермента синтеза эстрогенов in situ – ароматазу и 17β-гидроксистероиддегидрогеназу, тогда как в неизмененных клетках миометрия (ткань далее 2 см от капсулы миомы) подобная экспрессия отсутствует [28]. Тем не менее не выявлено связи между уровнем мРНК ароматазы и размером узла, объемом матки и возрастом пациенток. Лишь у пациенток старше 45 лет определена положительнвая корреляция между экспрессией ароматазы и возрастом.
Повышению концентрации эстрадиола в ткани способствует не только активация его синтеза, но и ингибирование процессов катаболизма. Катехол-0-метилтрансфераза (КОМТ) – фермент, осуществляющий конверсию катехолэстрогенов в их метиллированные производные. Концентрация катехолэстрогенов в миомном узле достоверно ниже, чем в здоровом миометрии. Существует несколько теорий о роли КОМТ и катехолэстрогенов в патофизиологии ЛММ. Во-первых, КОМТ в качестве субстрата использует 2-гидроксиэстрадиол, проявляющий антиэстрогенную активность. За счет уменьшения уровня эндогенных антиэстрогенов усиливается эстрогенное влияние. Другими словами, повышение активности КОМТ сопровождается гиперэстрогенией, способствующей прогрессированию миомы. В свою очередь продукт активности КОМТ – 2-метоксиэстрадиол – проявляет митогенное действие. Установлено, что катехолэстрогены (субстрат КОМТ) стимулируют образование гидроксильных радикалов, которые активируют тканевые металлопротеиназы, гидролизующие белки межклеточного матрикса. Именно межклеточный матрикс является важнейшим компонентом миоматозного узла. Поэтому активация КОМТ, приводящая к истощению запаса катехолэстрогенов, ведет к накоплению материала межклеточного матрикса [44].
Популяционный анализ показал, что у пациенток афроамериканской расы наряду с более высокой частотой встречаемости ЛММ активность КОМТ в миометрии значительно выше, чем у европейских женщин. Уровень КОМТ в миомном узле выше, чем в окружающем миометрии. Более того, активность КОМТ в узле коррелирует с размером узла. Не удивительно, что активность КОМТ находится под гормональным контролем в здоровой и опухолевой ткани, при этом эстрадиол подавляет экспрессию КОМТ, а прогестерон и глюкокортикоиды стимулируют ее. Следовательно, КОМТ – потенциальная мишень в лечении ЛММ, а применение антигестагенов или антиглюкокортикоидов будет сдерживать активность КОМТ в ЛММ и, следовательно, ее пролиферацию.
Накопление активного эстрогена в клетках миомы ведет к усилению экспрессии рецепторов эстрадиола (РЭ) и рецепторов прогестерона (РП), гиперчувствительности к гормонам и росту миомы. С этим согласуются и данные о более высокой способности клеток миомы к пролиферации в течение менструального цикла по сравнению с неизмененным миометрием.
Рецепторы половых стероидов в ЛММ
Уровень РЭ (обе изоформы) в ЛММ значительно выше, чем в аутентичной ткани [13]. При этом на циклические изменения половых гормонов в плазме крови больше отвечают РЭ-α, тогда как колебание концентрации РЭ-β значительно меньше. Однако в литературе имеются данные о том, что у польских женщин детородного возраста уровень РЭ одинаков в ЛММ и ГНТМ (при этом везде доминирует РЭ-α) [33], а также о том, что в отличие от окружающего миометрия в ткани миомы уровень РЭ остается статичным в течение всего менструального цикла (Shozu M. и соавт., 2000). Сравнительный анализ четырех этнических групп пациенток (афроамериканок, азиаток, испанок и представительниц белой расы) показал, что уровень РЭ-α как в ЛММ, так и в ГНТМ повышен у афроамериканок по сравнению с представительницами других рас. Повышенный уровень РЭ в узле определяет более высокую его чувствительность к эстрадиолу по сравнению с неизмененной тканью.
Содержание РП в миомных узлах также значительно выше, чем в неповрежденном миометрии и соответствует их уровню в эндометрии [7, 49], при этом количество РП в миоме в течение менструального цикла изменяется так же, как и в неизмененном миометрии [30]. Наибольшее количество РП в миометрии отмечается в середине цикла; в секреторную фазу их уровень уменьшается. Интересны данные о том, что в субмукозных узлах уровень РП может быть выше, чем в субсерозных [17]. Эти результаты согласуются с гипотезой, что лейомиомы имеют различную природу: субсерозные узлы развиваются из периваскулярных гладкомышечных фибробластов, а субмукозные узлы – из миометриальных клеток и по-разному реагируют на гестагенотерапию.
Многие исследователи отмечают повышение уровня обеих изоформ РП в ЛММ по сравнению с ГНТМ. При этом одни авторы обнаружили равномерное повышение, тогда как другие указывают на преимущественный рост уровня РП-А типа над РП-В типом в ЛММ [42]. Прогестероновые рецепторы не ограничиваются только ядерными вариантами РП-А и РП-В, описаны свойства по меньшей мере еще двух типов РП в клетке-мишени – мембраносвязанный и θ-тип. В доступной литературе крайне мало данных о рецепции половых стероидов на уровне плазматических мембран клеток миомы.
На кафедре молекулярной фармакологии и радиобиологии медико-биологического факультета (зав. – чл.-корр. РАМН, профессор Шимановский Н.Л.) и кафедре акушерства и гинекологии педиатрического факультета (зав. – академик РАМН, профессор Савельева Г.М.) РГМУ проведен сравнительный анализ не только цитозольной, но и плазмамембранной рецепции половых стероидов в тканях матки при миоме [8]. В результате показано, что миоматозная ткань отличается более высоким уровнем цитозольных рецепторов половых стероидов по сравнению с ГНТМ. При этом рецепция стероидов в плазматических мембранах клеток миомы характеризуется снижением числа связывающих мест для прогестерона при неизмененном связывании эстрадиола (ВmaxE2/BmaxP4 – в 2,5 раза выше во фракции плазматических мембран миомы по сравнению с ГНТМ). Установлено, что при ЛММ изменяется липидный матрикс мембран клеток миометрия, активность 5’-нуклеотидазы, фосфолипазы С, уровень цГМФ (циклического гуанозин-3,5-монофосфата). При этом наиболее информативным критерием развития гиперпластических процессов в миометрии является изменение плазмамембранной рецепции прогестерона [2].
Транскрипционные эффекты прогестерона широко известны, приведем лишь один пример: в клетках ЛММ прогестерон и синтетический гестаген R5020 стимулируют AKT-путь, обеспечивающий выживание и пролиферацию клеток [10, 47].
Таким образом, роль прогестерона в росте ЛММ не вызывает сомнений. Поэтому не удивительно, что антигестагены: мифепристон, азоприснил и CDB-2914 (17α-acetoxy-11β-[4-N,Ndimethylaminophenyl]-19-norpregna4,9-diene-3,20-dione), эффективно уменьшают объем миомных узлов.
Агонисты гонадолиберинов также уменьшают размер узлов, однако, если одновременно с аГРГ использовать гестагены, снижения объема ЛММ не происходит.
Половые стероиды и митотическая активность миометрия
Особого внимания заслуживает тот факт, что наиболее высокий митотический индекс в клетках миомы отмечается в лютеиновую фазу менструального цикла, что поддерживает гипотезу об этиопатогенетической роли прогестерона в развитии миомы матки. Существуют клинические, паталогоанатомические и молекулярно-биохимические доказательства того, что прогестерон и активация его рецепторов стимулируют пролиферацию клеток миомы. Экспрессия маркера пролиферативной активности клеток – антигена Ki-67 – в ЛММ снижена в фолликулярной и повышена в лютеиновой фазах менструального цикла и при беременности, т. е. в периоды повышенной концентрации прогестерона. Увеличение митотической активности показано в миомных узлах у женщин, получавших гестагенную терапию (МПА). Пролиферативная активность ЛММ у пациенток в постменопаузе значительно ниже, чем в пременопаузе. В постклимактерическом периоде при условии отсутствия заместительной гормональной терапии деления клеток в узле вообще не происходит.
Приобретение лейомиоматозного фенотипа клетками отражается на их чувствительности к половым стероидам. Так, in vitro добавление эстрадиола и прогестерона (Р4) к культуре ЛММ вызывает повышение митотического индекса, в то время как в неизмененных клетках только эстрадиол вызывает пролиферативный ответ (не прогестерон!!!) [38].
Таким образом, повышенная экспрессия рецепторов половых стероидов в ткани ЛММ неопровержимо свидетельствует об участии этих гормонов в промоции узлов миомы. Более того, успешная антигормональная терапия ЛММ не оставляет сомнений, что наиважнейшим уровнем внутриклеточной
регуляции пролиферации клеток являются именно стероидные рецепторы. Активация этих рецепторов занимает наиболее высокое положение в иерархической лестнице регуляции деления клеток. Все последующие за активацией события являются вторичными, и поэтому прерывание сигнала в узловых точках патологического каскада (стероидных рецепторах) неминуемо влечет за собой торможение нисходящих путей и в конечном итоге – остановку пролиферации миомных клеток. Конечно, нельзя умалять значение нисходящих звеньев регуляции деления клеток (факторов роста, апоптоза, ангиогенеза) в качестве мишеней для терапии миомы матки, особенно с учетом того факта, что антигормональная терапия хоть и успешна, но ограничена в детородном возрасте, т. к. плохо сочетается с репродуктивными планами пациенток.
регуляции пролиферации клеток являются именно стероидные рецепторы. Активация этих рецепторов занимает наиболее высокое положение в иерархической лестнице регуляции деления клеток. Все последующие за активацией события являются вторичными, и поэтому прерывание сигнала в узловых точках патологического каскада (стероидных рецепторах) неминуемо влечет за собой торможение нисходящих путей и в конечном итоге – остановку пролиферации миомных клеток. Конечно, нельзя умалять значение нисходящих звеньев регуляции деления клеток (факторов роста, апоптоза, ангиогенеза) в качестве мишеней для терапии миомы матки, особенно с учетом того факта, что антигормональная терапия хоть и успешна, но ограничена в детородном возрасте, т. к. плохо сочетается с репродуктивными планами пациенток.
Основными эффектами половых стероидов в клетках лейомиомы являются стимуляция экспрессии тканевых факторов роста и их рецепторов, антиапоптотических факторов, регуляторов клеточного цикла и ингибирование синтеза проапоптотических белков.
Половые стероиды и пептидные регуляторные факторы
Отдельные тканевые факторы роста, апоптоза и клеточного цикла находятся под контролем половых гормонов. Эти факторы промотируют митогенное и фиброгенное действия в ткани миомы. Основные миом-ассоциированные факторы роста: трансформирующий (ТФР, TGF – transforming growth factor), основной фактор роста фибробластов (оФРФ, bFGF – basic fibroblast growth factor), эпидермальный (ЭФР, EGF – epidermal growth factor), тромбоцитарный ростовой фактор (ТРФ, PDGF – platelet-derived growth factor) сосудистый эндотелиальный (СЭФР,VEGF – vascular endothelial
growth factor), инсулиноподобные (ИПФР, IGF – insulin-growth factor) и пролактин [23]. Многие из этих ростовых факторов оверэкспрессированы в ЛММ и а) увеличивают пролиферацию утеромиоцитов, б) увеличивают синтезы ДНК, в) стимулируют синтезы компонентов межклеточного матрикса, г) промотируют митогенез, д) усиливают ангиогенез. Похоже, и другие миомассоциированные факторы роста будут открыты, и остается ждать, какой из них будет признан ведущим [55].
growth factor), инсулиноподобные (ИПФР, IGF – insulin-growth factor) и пролактин [23]. Многие из этих ростовых факторов оверэкспрессированы в ЛММ и а) увеличивают пролиферацию утеромиоцитов, б) увеличивают синтезы ДНК, в) стимулируют синтезы компонентов межклеточного матрикса, г) промотируют митогенез, д) усиливают ангиогенез. Похоже, и другие миомассоциированные факторы роста будут открыты, и остается ждать, какой из них будет признан ведущим [55].
Трансформирующий фактор роста (ТФР) регулирует рост клеток, вовлечен в процессы воспаления, апоптоза и ремоделирования ткани, играет принципиальную роль в формировании межклеточного матрикса. В клетках ЛММ нарушена экспрессия некоторых изоформ данного цитокина и повышен уровень Smads, которые являются компонентами TФР-β-сигнального пути (рис. 1). Концентрация TФР-β и его рецепторов в ЛММ выше, чем в аутентичном миометрии, и достигает максимума в лютеиновую фазу менструального цикла [55]. Эстрадиол и прогестерон потенцируют действие TФР-β (рис. 1).
Рисунок 1. Половые стероиды и пути проведения сигнала ТФР-в в клетке-мишени.
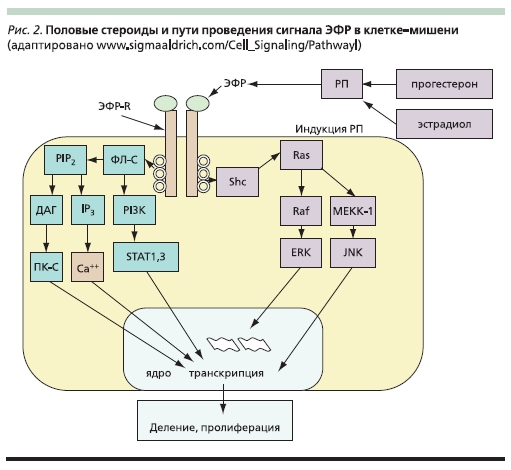
Рисунок 2. Половые стероиды и пути проведения сигнала ЭФР в клетке-мишени.
Эпидермальный фактор роста (ЭФР) оказывает митогенный эффект на ряд репродуктивных тканей. В культуре клеток ЛММ прогестерон повышает его уровень, а эстрадиол оказывает потенцирующее действие (рис. 2). Отсутствуют данные о флуктуациях экспрессии ЭФР и его рецепторов в тканях матки в течение менструального цикла. Данные о количестве ЭФР в фиброиде противоречивы: от отсутствия различий с нормальной тканью до повышенной экспрессии в фиброиде или прилегающем миометрии. Применение в модельной системе селективного антагониста рецептора ЭФР вызывает подавление пролиферации клеток ЛММ, которое не снимается половыми гормонами. Интересны данные о повышенной чувствительности рецепторов ЭФР в ткани фиброида по сравнению с ГНТМ [45].
Рисунок 3. Половые стероиды и пути проведения сигнала ТФР-в в клетке-мишени.
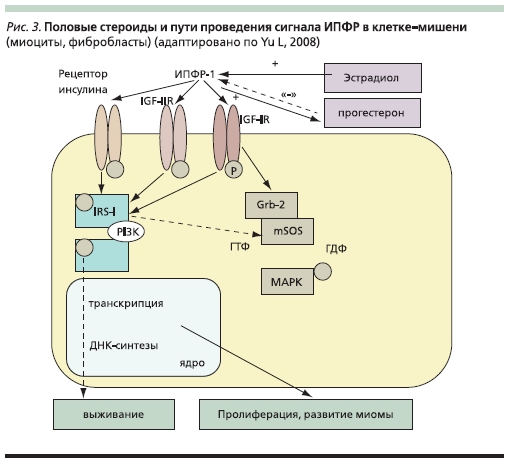
Инсулиноподобный фактор роста. ИПФР-1 – структурный аналог проинсулина – обеспечивает пролиферацию, дифференцировку и выживаемость клеток. Рецепторы ИПФР обладают тирозинкиназной активностью, в качестве вторичных посредников впередаче сигнала в клетку используют адапторы – IRS-I/Shc, которые через IRS/PI3K/AKT в свою очередь обеспечивают выживание клетки и через Shc/Ras/Crb2/MAP-киназы – клеточную пролиферацию (рис. 3). Многие авторы настаивают на ведущей роли данного ростового фактора в пролиферации миоцитов, т. к. при его недостаточности (экспериментально) не проявляют своей митогенной активности ни ЭФР, ни ТРФ, ни инсулин. В свою очередь для проявления митогенного эффекта ИПФР не нужны дополнительные факторы. Экспрессия ИПФР-1 в ткани ЛММ коррелирует с уровнем эстрадиола (промотор гена данного фактора содержит место связывания для РЭ) и в три раза выше по сравнению с прилегающим миометрием. Оверэкспрессия самого фактора и его рецептора обнаружена как в гладкомышечных клетках, так и в фибробластах ЛММ. ИПФР и его сигнальный путь нарушены в трети изученных фиброидов. Выявлена положительная корреляция между оверэкспрессией ИПФР-1 и размерами миомы [46]. Прогестерон демонстрирует сложное действие на ЛММ, которое в конечном итоге оборачивается митогенным эффектом [59].
Ангиогенез. Скрининг экспрессии генов (более 10,5 тыс.) в миомном узле и аутентичном миометрии, миоцитах и эндотелии был проведен Weston G. (2003) с акцентом на гены, продукты которых участвуют в ремоделировании сосудов [56]. В результате идентифицированы двадцать пять генов с
различиями экспрессии. Среди них – ИПФР-2, рецептор эндотелина, фактор роста соединительной ткани (ФРСТ, CTGF – connective tissue growth factor), богатый цистеином ангиогенный индуктор 61 (CYR61) и коллаген 4α2 (COL4A2). Фактор роста соединительной ткани и CYR61 кроме способности стимулировать ангиогенез регулируют клеточную адгезию, миграцию и продукцию компонентов внеклеточного матрикса; участвуют в процессах воспаления и репарации тканей. Кроме того, их оверэкспрессия отмечена в злокачественных опухолях молочной железы, в тканях с бурно протекающим фиброзом (склеродермия, келоид и фиброз легких). В итоге ткань миомы демонстрирует антиангиогенный профиль по сравнению с прилежащим миометрием, что может быть приемлемым объяснением сниженной плотности капиллярной сети в узлах. И действительно, во всех исследованиях показано, что в ткани миомы редуцирован кровоток по сравнению с прилежащим миометрием и миометрием матки без миомы [24]. Увеличение плотности сосудов отмечается в перифиброидной области прилежащего миометрия с большим количеством мелких сосудов, проникающих в ткань опухоли. После менопаузы плотность капиллярной сети миометрия увеличивается, а в фиброиде не изменяется, т. е. усугубляется разница кровоснабжения ткани в менопаузе – естественной модели регресса миомы.
различиями экспрессии. Среди них – ИПФР-2, рецептор эндотелина, фактор роста соединительной ткани (ФРСТ, CTGF – connective tissue growth factor), богатый цистеином ангиогенный индуктор 61 (CYR61) и коллаген 4α2 (COL4A2). Фактор роста соединительной ткани и CYR61 кроме способности стимулировать ангиогенез регулируют клеточную адгезию, миграцию и продукцию компонентов внеклеточного матрикса; участвуют в процессах воспаления и репарации тканей. Кроме того, их оверэкспрессия отмечена в злокачественных опухолях молочной железы, в тканях с бурно протекающим фиброзом (склеродермия, келоид и фиброз легких). В итоге ткань миомы демонстрирует антиангиогенный профиль по сравнению с прилежащим миометрием, что может быть приемлемым объяснением сниженной плотности капиллярной сети в узлах. И действительно, во всех исследованиях показано, что в ткани миомы редуцирован кровоток по сравнению с прилежащим миометрием и миометрием матки без миомы [24]. Увеличение плотности сосудов отмечается в перифиброидной области прилежащего миометрия с большим количеством мелких сосудов, проникающих в ткань опухоли. После менопаузы плотность капиллярной сети миометрия увеличивается, а в фиброиде не изменяется, т. е. усугубляется разница кровоснабжения ткани в менопаузе – естественной модели регресса миомы.
Сосудистый эзндотелиальный фактор роста (СЭФР). Экспрессия СЭФР-A в ЛММ выше, чем в здоровом миометрии. Эффект СЭФР-A сводится к стимуляции пролиферации и миграции эндотелиальных клеток и вазодилатации за счет высвобождения вазоактивных субстанций – нитроксида и простациклина. Эстрадиол и прогестерон стимулируют экспрессию мРНК СЭФР-А и СЭФР-В в культуре клеток ЛММ и здорового миометрия [57]. В промоторной области гена СЭФР-А выявлен участок, потенциально способный связывать РП (прямая стероидная регуляция экспрессии).
Адреномедуллин – наряду с СЭФР регулирует ангиогенез, в т. ч. в ткани ЛММ. Адреномедуллин относится к суперсемейству пептидов кальцитонина. Экспрессия адреномедуллина тесно связана с плотностью сосудов и пролиферацией эндотелиальных клеток в миометрии и фибромиоме. Aдреномедуллин участвует в карциногенезе и прогрессировании многих опухолей через стимуляцию пролиферации, ангиогенеза и ингибирование апоптоза. Прогестерон стимулирует экспрессию адреномедуллина и его рецепторов в тканях матки (Xu Q, 2006).
Фактор некроза опухоли (ФНО, TNF – tumor necrosis factor) – секретируется не только активированными макрофагами, но и многими типами клеток женского репродуктивного тракта, ингибирует пролиферацию и стимулирует апоптоз многих клеток [32]. Экспрессия ФНО в ЛММ выше, чем в окружающем миометрии с максимальными значениями в пролиферативной фазе. Уровень ФНО в аутентичном миометрии не изменяется в ходе полового цикла. In vitro прогестерон (3,18 × 10-7 моль/л) ингибирует экспрессию ФНО в культуре клеток ЛММ, эстрадиол (3,67 × 10-8 моль/л) не влияет на уровень ФНО. Таким образом, одним из механизмов пролиферативного эффекта прогестерона на клетки миомы может быть подавление экспрессии ФНО (Kurachi O., 2001). Факторы апоптоза. В ЛММ выявлена оверэкспрессия фактора апоптоза bcl-2 по сравнению с ГНТМ. В отдельных клетках оверэкспрессия bcl-2 может промотировать клеточную репликацию, уменьшая потребность последних в ростовых факторах. Показано, что половые стероиды, особенно прогестерон, стимулируют экспрессию bcl-2 в ЛММ – промоторная область гена bcl-2 содержит участки, содержащие специфические гормоночувствительные элементы и потому способна связывать РП, что в конечном итоге ведет к активации экспрессии bcl-2 [58]. Не удивительно поэтому, что экспрессия фактора апоптоза bcl-2 в ткани ЛММ и ГНТМ достигает максимальных значений в лютеиновую фазу менструального цикла. Интересно, что эстрадиол снижает уровень bcl-2 в указанных тканях [36]. In vitro прогестерон подавляет апоптоз и стимулирует пролиферацию миоцитов ЛММ, тогда как антагонисты прогестерона ингибируют пролиферацию и стимулируют апоптоз в ткани ЛММ [36]. В то же время при беременности прогестерон в миометрии поддерживает уровень проапоптотического фермента каспазы-3, тем самым обеспечивая пролиферативный баланс ткани [29].
Таким образом, нарушение баланса между процессами пролиферации и апоптоза занимает одну из ключевых позиций в патобиозе миомы матки. Особого внимания заслуживает тот факт, что в отличие от нормальной ткани в лейомиоме нарушен контроль апоптоза половыми стероидами, что открывает возможности поиска перспективных лекарственных препаратов, способных взять под контроль апоптоз в ЛММ.
Регуляторы клеточного цикла. Пролиферация клеток регулируется механизмами контроля клеточного цикла, включающего набор циклинзависимых киназ (CDK – cyclin-dependent kinases, серин/треонин-протеинкиназы) вкупе с их активаторами (циклинами) и ингибиторами. Функция CDK – фосфорилирование протеинов, принимающих участие в митозе, таких, например, как входящий в состав хроматина гистон H1, ламин (компонент цитоскелета), факторы транскрипции, белки митотического веретена и ряд ферментов. Фосфорилирование перечисленных белков запускает процесс митоза. После завершения митоза циклин подвергается протеолизу. Далее наступает очередь фосфатаз, которые дефосфорилируют белки, принимавшие участие в митозе, после чего клетка возвращается в состояние интерфазы. CDK активны только в комплексе с циклинами. Циклин непрерывно синтезируется в интерфазе клеточного цикла до достижения максимальной концентрации в фазе М, когда запускается весь каскад фосфорилирования белков, катализируемый CDK. К окончанию митоза циклин быстро разрушается протеиназами. Действие циклинов и ассоциированных с ними киназ блокируется ингибиторами циклин-зависимых киназ – CKI (cyclin dependent kinase inhibitors), представленных двумя семействами Ink4 (р16, р15, р18 – селективные ингибиторы CDK4/6) и Cip/Kip (р21, р27, р57 (росковитин – фармацевтический препарат) – универсальные ингибиторы CDK) (Chen B., 2005).
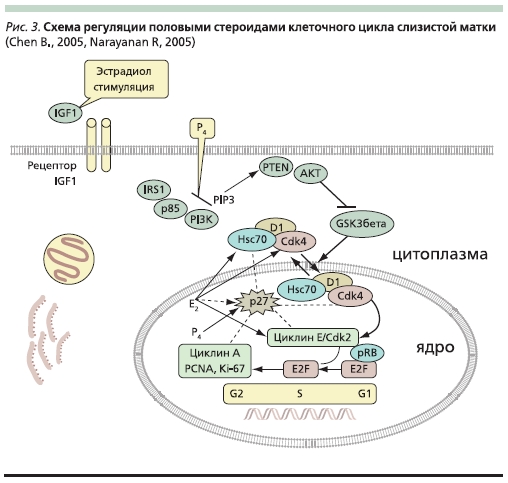
Рисунок 4. Схема регуляции половыми стероидами клеточного цикла слизистой матки.
Ключевой точкой влияния половых стероидов является циклин D1, вернее его транспорт в ядро (рис. 4). Известно, что принципиальной в этом транспорте циклина является активность гликогенсинтазы киназы-3β (GSK-3β – glycogen synthase kinase-3β), стимулирующая ее вход в ядро, фосфорилируя молекулу циклина D1. Эстрадиол прямо через PI3-киназу – АКТ – GSK-3β регулирует клеточный цикл (эпителиальные клетки эндометрия). Кроме того, эстрадиол в раннюю G1-стадию увеличивает активность CDK4 и CDK2, стимулирует экспрессию циклина D1, а также снижает уровень ингибиторов CDK. Уровень циклина D1 в ткани ЛММ у пациенток во все стадии менструального цикла достоверно выше по сравнению с окружающим миометрием [31].
Концентрация р27 – ингибитора CDK – варьируется в клетке в зависимости от фазы клеточного цикла. Уровень ингибиторов циклинзависимых киназ резко снижен во многих злокачественных опухолях кожи, печени, молочной железы, простаты, эндометрия и др. Причем существует тесная корреляция между уровнем ингибитора (р27) и степенью агрессивности опухолевого процесса. Экспрессия р27 находится под гормональным контролем; так, эстрадиол подавляет ее, а прогестерон стимулирует в железистых клетках эндометрия (доступных данных по миоме нет). Несмотря на исторически сложившееся представление о р27 как об антипролиферативном факторе, в последнее время стали появляться сообщения о возможной комбинации про- и антиапоптотических свойств этого протеина в зависимости от клеточного и тканевого контекстов [48]. Экспрессия р27 в ткани миомы ниже, чем в окружающем миометрии.
Но более интересными нам представляются сведения о регуляции циклинами транскрипционной активности рецепторов стероидных гормонов. В частности, циклин A/CDK2 выступает в роли облигатного коактиватора РП, который взаимодействует с циклином А и рекрутирует циклин A/CDK2 к прогестинувствительным промоторам, стимулируя транскрипцию прогестерон-зависимых генов. По-видимому, такое взаимодействие циклинов и РП необходимо для трасдукции гормонального сигнала, т. к. подавление CDK2 ведет к снижению транскрипционной активности гормонрецепторного комплекса [41]. В свою очередь циклин D1 взаимодействует с лиганд-связывающим доменом РЭ и стимулирует его трансактивацию, а циклин А – нет [54]. Остальные рецепторы суперсемейства также не остаются без внимания циклинов; так, циклин Е работает как коактиватор андрогенового рецептора, а циклин D1 ингибирует андрогенную активность.
Таким образом, взаимодействие рецепторов половых стероидов и системы регуляции клеточного цикла является многокомпонентным и разноуровневым. Если говорить об иерархии внутриклеточных сигналов в ткани миомы, то приходится признать, что при объективной значимости клеточного контекста (в данном случае речь идет о циклинах и зависимых киназах как корегуляторах транскрипции) ключевая роль в регуляции пролиферации миоцитов принадлежит обоим стероидам.
Таким образом, контроль роста ЛММ изучен не полностью, но наиболее важные его детерминанты определены – это половые стероиды, тканевые факторы роста и стероидрегулируемые протоонкогены [26].
Образование и рост лейомиомы связаны со сложным взаимодействием в миометрии и опухоли стероидных гормонов и их рецепторов. Половые стероиды регулируют рост ЛММ посредством комплекса факторов роста и цитокинов, продуцируемых гладкомышечными клетками и фибробласта-
ми, влияющими на процессы пролиферации, апоптоза и ангиогенеза [24].
ми, влияющими на процессы пролиферации, апоптоза и ангиогенеза [24].
В целом пролиферативная активность миометрия при миоме значительно отличается от ГНТМ как по уровню выраженности, так и по контролируемости процесса. Растет число биохимических, гистологических, клинических и фармакологических доказательств о ключевой роли прогестерона в росте и развитии ЛММ.
Некоторые из них:
1. Повышенный уровень РП (обеих изоформ) в ткани ЛММ по сравнению с аутологичным миометрием. Выявлена тесная положительная корреляция между интенсивностью роста узлов и уровнем в них РП (Ichimura T., 1998).
2. In vitro прогестерон подавляет апоптоз и стимулирует пролиферацию миоцитов ЛММ, тогда как антагонисты прогестерона ингибируют пролиферацию и стимулируют апоптоз в ткани ЛММ.
3. Активация митозов в ткани миомы в прогестерон-доминантной фазе менструального цикла.
4. Увеличение митотической активности ЛММ у пациенток, получающих терапию гестагенами.
5. В культуре клеток прогестерон повышает уровень протеинов, включенных в клеточную пролиферацию; дифференцировочный статус клеток ЛММ отрицательно регулируется прогестероном и положительно – эстрадиолом (Okolo S., 2009).
6. Синтетические гестагены (МПА и норэтинодрон) при использовании их в качестве поддерживающей терапии на фоне введения аГРГ снижают или даже блокируют уменьшение объема миоматозных узлов.
1. Повышенный уровень РП (обеих изоформ) в ткани ЛММ по сравнению с аутологичным миометрием. Выявлена тесная положительная корреляция между интенсивностью роста узлов и уровнем в них РП (Ichimura T., 1998).
2. In vitro прогестерон подавляет апоптоз и стимулирует пролиферацию миоцитов ЛММ, тогда как антагонисты прогестерона ингибируют пролиферацию и стимулируют апоптоз в ткани ЛММ.
3. Активация митозов в ткани миомы в прогестерон-доминантной фазе менструального цикла.
4. Увеличение митотической активности ЛММ у пациенток, получающих терапию гестагенами.
5. В культуре клеток прогестерон повышает уровень протеинов, включенных в клеточную пролиферацию; дифференцировочный статус клеток ЛММ отрицательно регулируется прогестероном и положительно – эстрадиолом (Okolo S., 2009).
6. Синтетические гестагены (МПА и норэтинодрон) при использовании их в качестве поддерживающей терапии на фоне введения аГРГ снижают или даже блокируют уменьшение объема миоматозных узлов.
Медикаментозное лечение миомы матки
ЛММ практически в половине установленных случаев протекают бессимптомно. В этой ситуации лечение не показано. Появление симптомов сигнализирует о необходимости начала лечения. Наиболее частые симптомы – тяжелые или болезненные менструальные кровотечения, которые могут вызывать анемию, межменструальные кровотечения, боль и чувство тяжести внизу живота, – все это по отдельности или в совокупности значительно ухудшает качество жизни пациенток.
Патогенетически обоснованной концепцией лечения ЛММ является комбинированное воздействие – хирургическое и медикаментозное. Поэтому, несмотря на появление новых оперативных технологий, медикаментознаяя терапия не утратила своей значимости. Консервативное лечение должно быть комплексным и направлено на различные звенья патологического процесса с учетом выявленных нарушений. Базовыми препаратами патогенетического лечения ЛММ являются препараты с антигормональной активностью. Это вполне закономерно с учетом ведущей роли половых стероидов в патогенезе данной нозологической формы заболевания. Снизить эффект эндогенных гормонов на ткань-мишень можно несколькими способами. Во-первых, это центральная блокада секреции гонадорелина (агонисты и антагонисты ГРГ) со снижением продукции гонадотропинов (лютеинизирующего гормона – ЛГ, фолликулостимулирующего гормона – ФСГ) и угнетением функции яичников. В этом случае снижается уровень одновременно и прогестерона, и эстрадиола; эффективно снижается объем миомных узлов и уходит негативная симптоматика.
Второй подход – дифференцированная блокада гормональных сигналов (эстрадиола или прогестерона). Антиэстрогенная стратегия включает использование: 1) гестагенов и андрогенов, проявляющих антагонизм к значительному числу эстрогензависимых процессов в разных тканях; 2) собственно антиэстрогенов и селективных модуляторов рецепторов эстрадиола (только в комбинации с аГРГ); 3) ингибиторов синтеза активных эстрогенов. Отсутствие показаний к монотерапии ЛММ антиэстрогенами еще раз подчеркивает ведущее значение проведения прогестеронового сигнала в клеткемишени для ее пролиферации.
Исторически значительно позже появились доказательства участия прогестерона в патогенезе миомы, поэтому эта область является более молодой, но чрезвычайно активно развивающейся. Антигестагенная стратегия подразумевает использование известных и новых антигестагенов, а также
селективных модуляторов РП (мифепристона, азоприснила). Сведения о текущей эффективности медикаментозной терапии ЛММ опубликованы в нескольких аналитических работах [34, 51].
селективных модуляторов РП (мифепристона, азоприснила). Сведения о текущей эффективности медикаментозной терапии ЛММ опубликованы в нескольких аналитических работах [34, 51].
Опираясь на критерии доказательной медицины, авторы суммарно пришли к следующим выводам:
1. Агонисты ГРГ наиболее широко используются в лекарственной терапии ЛММ. Они действительно вызывают регресс миомы, уменьшают объемы матки, миомы, кровотечения, но не могут использоваться длительно в силу значительной гипоэстрогении. Есть отдельные свидетельства, что присоединенная терапия (гестаген, тиболон, комбинация эстроген/гестаген, ралоксифен) может уменьшать симптомы гипоэстрогении; но нет достаточного числа исследований, соответствующих современным требованиям доказательной медицины. Применение аГРГ, по крайней мере в течение 3 месяцев перед оперативным вмешательством по поводу ЛММ, повышает концентрацию гемоглобина и гематокрит, уменьшает выраженность миомассоциированной симптоматики. Ограничение длительности возможного применения препаратов предопределяет ограничение в показаниях: 1) пациенткам в пременопаузальном возрасте (в расчете на то, что после окончания лечения функция яичников не восстановится) и 2) для подготовки к операции.
2. Антагонисты ГРГ не вызывают начального выброса ЛГ и ФСГ, характерного для агонистов ГРГ, но никаких дополнительных преимуществ перед аналогами, похоже, не имеют.
3. Антагонисты РЭ. Представлено слабое свидетельство того, что ралоксифен может быть полезным для пациенток позднего пременопаузального возраста (с низким уровнем системного эстрадиола). Ралоксифен доказанно эффективно вызывает регресс миомы только у пациенток в постменопаузальном возрасте. Аналогично селективным эстрогенным рецепторным модуляторам ингибиторы ароматазы доказанно эффективны только у женщин после менопаузы, при этом препараты проявляют существенные долгосрочные побочные эффекты; опять же опыт их использования при ЛММ ограничен. Актуальность медикаментозной терапии ЛММ у пациенток с угасшей функцией яичников не вели-ка, т. к. в этом возрастном периоде подавляющее большинство ЛММ имеют обратное разитие или торможение процесса без дополнительных вмешательств.
4. Левоноргестрел в составе внутриматочной системы очевидно и значимо сокращает ежемесячные кровопотери у женщин с ЛММ, но до настоящего времени не было проведено его клинических испытаний, соответствующих принципам доказательной медицины, у пациенток с ЛММ.
5. Антигестагены. Антагонисты прогестерона мифепристон и азоприснил вселяют большие надежды. Они показали многообещающие результаты по эффективности и безопасности применения, что гарантирует их дальнейшее исследование и применение. Главное их преимущество – высокий эффект в отсутствие грозных побочных эффектов. Однако они, как и другие гормональные методы лечения, отодвигают реализацию репродуктивных планов пациенток.
Мифепристон
Основным достижением репродуктивной эндокринологии 1980-х гг. был синтез мифепристона (RU486) – высокоаффинного антигестагена. В дозе 200–800 мг с мизопростолом или без него мифепристон надежно вызывает прерывание беременности во все сроки гестации. Столь востребованный эффект затмил на некоторое время другие возможности использования мифепристона. В дальнейшем изучение мифепристона в ведущих клиниках мира показало, что препарат может применяться в качестве лекарственного средства для экстренной контрацепции, для расширения шейки матки при доношенной беременности в процессе подготовки к родам, а также для лечения ЛММ. Сейчас проводятся широкие клинические испытания антидепрессивных свойств препарата.
В 1993 г. завершено первое исследование мифепристона в лечении миомы матки [39]. Мифепристон назначали 10 женщинам в возрасте 18–45 лет с регулярными менструациями и симптомной миомой матки в суточной дозе 50 мг в течение 3 месяце, начиная с 1–3-го дня менструального цикла. Через 8 недель лечения наблюдалось достоверное снижение объема миомы. Среднее уменьшение объема составило 22 % через 4 недели, 39 % – через 8 и 49 % – через 12 недель лечения. Темп был сравним с таковым при применении аГРГ в течение 6 месяцев [18]. Основным механизмом действия антигестагенов является компетентное ингибирование связывания прогестерона с РП и торможение прогестерониндуцированной транскрипции генов-мишеней. Мифепристон также проявляет антагонизм с глюкокортикостероидами и андрогенами. Препарат активен при пероральном применении. Максимальная концентрация достигается через 1,5 часа. Абсолютная биодоступность составляет 69 %. В плазме крови препарат на 98 % связывается с белками: альбумином и кислым α1-гликопротеином. После фазы распределения выведение происходит сначала медленно, концентрация уменьшается в два раза между 12–72 часами. Период полувыведения составляет 18 часов. Мифепристон уменьшает число РП в ЛММ и ГНТМ, поддерживает гормональный фон, соответствующий ранней фолликулярной фазе, прямо влияет на сосуды матки, уменьшая кровоток. Более высокие дозы мифепристона (25–50 мг) способны активировать гипоталамо-гипофизарную ось, что вызывает повышение плазменного адренокортикотропного гормона и как следствие – увеличивает не только плазменный кортизол, но и надпочечниковый андроген и эстрадиол. Это в свою очередь способствует гиперплазии эндометрия. Важно, что пока не отмечено ни одного случая появления эндометриальной карциномы при длительном применении мифепристона. Кроме того, мифепристон проявляет свойства антиоксиданта, ингибирует продукцию гистиоцитами гликоделина, индуцированную лизофосфатидной кислотой [48].
В целом результаты клинических испытаний мифепристона в терапии ЛММ сводятся к следующему:
• Дозы: эффективны дозы мифепристона от 5 до 50 мг/сут перорально. Возможно применение мифепристона по 50 мг через день, 10 мг вагинально. Эффект дозозависим (Bagaria M., 2008).
• Сроки: показана безопасность применения мифепристона в течение 3–12 месяцев.
• Особенности приема препарата: мифепристон назначают с 1–3-го дня менструального цикла, т. е. в ранней фолликулярной фазе до формирования доминирующего фолликула. Применение мифепристона в позднюю фолликулярную фазу приводит к коллапсу доминантного фолликула и прорывному кровотечению.
• Эффективность: мифепристон вызывает снижение объема матки на 27–49 % и миомных узлов – на 26–74 %; аменорею – у 40–70–100 % (5–10–100 мг); уменьшает объем менструальной кровопотери (Bagaria M., 2008); повышает уровень гемоглобина (50 мг через день 3 месяца) [22]; уменьшает проявления дисменореи и боли в тазовой области у 75 % пациенток уже через 2 месяца лечения. Результаты исследования продемонстрировали широкие вариации индивидуального ответа пациенток на лечение. Через полгода после завершения лечения у части пациенток (менее 20 %) наблюдалось увеличение объема матки, при этом максимальные значения размеров органа было на 42 % ниже, чем исходный уровень [21]. Сокращение размера ЛММ происходило без параллельного уменьшения кровоснабжения матки. У всех пациенток через 2–6 недель после прекращения лечения восстанавился менструальный цикл. В целом было отмечено значительное повышение качества жизни пациенток за счет уменьшения негативной симптоматики (De Leo и соавт., 2002).
• Безопасность: препарат хорошо переносится, доказательством чему служит практическое отсутствие пациенток, вышедших из исследований по причине побочных эффектов. Однако у 63 % пациенток в группе мифепристона (10 мг в течение года) выявлена простая гиперплазия эндометрия (в группе плацебо – 0), при этом ни одного случая атипии не было обнаружено [11]. Побочные эффекты включали вазомоторные симптомы (38 %), простую гиперплазию эндометрия (28 %), временное повышение уровня трансаминаз (4 %) [53]. Вызванные препаратом изменения эндометрия были обратимыми. Eisinger S.H. и соавт. (2005), используя две дозы (5 и 10 мг), обнаружили гиперплазию эндометрия (без атипии) только в группе, получавшей 10 мг: у 5 (13,9 %) из 36 пациенток – через шесть месяцев и только у одной (4,8 %) из 21 – в конце одного года лечения [21]. Возможно, что регресс эндометриальной гиперплазии произошел за счет возобновления менструаций при продлении
терапии.
• Дозы: эффективны дозы мифепристона от 5 до 50 мг/сут перорально. Возможно применение мифепристона по 50 мг через день, 10 мг вагинально. Эффект дозозависим (Bagaria M., 2008).
• Сроки: показана безопасность применения мифепристона в течение 3–12 месяцев.
• Особенности приема препарата: мифепристон назначают с 1–3-го дня менструального цикла, т. е. в ранней фолликулярной фазе до формирования доминирующего фолликула. Применение мифепристона в позднюю фолликулярную фазу приводит к коллапсу доминантного фолликула и прорывному кровотечению.
• Эффективность: мифепристон вызывает снижение объема матки на 27–49 % и миомных узлов – на 26–74 %; аменорею – у 40–70–100 % (5–10–100 мг); уменьшает объем менструальной кровопотери (Bagaria M., 2008); повышает уровень гемоглобина (50 мг через день 3 месяца) [22]; уменьшает проявления дисменореи и боли в тазовой области у 75 % пациенток уже через 2 месяца лечения. Результаты исследования продемонстрировали широкие вариации индивидуального ответа пациенток на лечение. Через полгода после завершения лечения у части пациенток (менее 20 %) наблюдалось увеличение объема матки, при этом максимальные значения размеров органа было на 42 % ниже, чем исходный уровень [21]. Сокращение размера ЛММ происходило без параллельного уменьшения кровоснабжения матки. У всех пациенток через 2–6 недель после прекращения лечения восстанавился менструальный цикл. В целом было отмечено значительное повышение качества жизни пациенток за счет уменьшения негативной симптоматики (De Leo и соавт., 2002).
• Безопасность: препарат хорошо переносится, доказательством чему служит практическое отсутствие пациенток, вышедших из исследований по причине побочных эффектов. Однако у 63 % пациенток в группе мифепристона (10 мг в течение года) выявлена простая гиперплазия эндометрия (в группе плацебо – 0), при этом ни одного случая атипии не было обнаружено [11]. Побочные эффекты включали вазомоторные симптомы (38 %), простую гиперплазию эндометрия (28 %), временное повышение уровня трансаминаз (4 %) [53]. Вызванные препаратом изменения эндометрия были обратимыми. Eisinger S.H. и соавт. (2005), используя две дозы (5 и 10 мг), обнаружили гиперплазию эндометрия (без атипии) только в группе, получавшей 10 мг: у 5 (13,9 %) из 36 пациенток – через шесть месяцев и только у одной (4,8 %) из 21 – в конце одного года лечения [21]. Возможно, что регресс эндометриальной гиперплазии произошел за счет возобновления менструаций при продлении
терапии.
В РФ были проведены клинические испытания препарата Мифепристон (МИР-ФАРМ ЗАО, Россия) по показанию лечение лейомиомы матки. Исследования проводили на базе Научного центра акушерства, гинекологии и перинатологии РАМН, а также кафедры акушерства и гинекологии ММА им. И.М. Сеченова на основании решения Фармакологического комитета МЗ РФ, протокол № 11/5 от 17.10.2002, и разрешения на проведение клинических испытаний Департамента госконтроля лекарственных средств и медицинской техники МЗ РФ № 280 от 26.12.2002. Целью клинических испытаний явилось определение эффективности мифепристона в монотерапии при использовании его для лечения ЛММ размером до 12 недель беременности [6].
В исследование вошли 60 пациенток в возрасте от 25 до 45 лет с ЛММ с интерстициальным и субсерозным расположением миоматозных узлов и отсутствием гиперпластического процесса в эндометрии. Из них 30 пациенток получали препарат в дозе 50 мг ежедневно перорально в течение 3 месяцев, а 30 пациенток получали препарат в дозе 25 мг ежедневно перорально также в течение 3 месяцев. У всех пациенток лечение начиналось со второго дня менструального цикла.
Мифепристон назначали с учетом имеющихся противопоказаний к его применению:
• наличие в анамнезе повышенной чувствительности к мифепристону;
• надпочечниковая недостаточность и длительная глюкокортикостероидная терапия;
• острая или хроническая почечная и/или печеночная недостаточность;
• порфирия;
• анемия;
• нарушения гемостаза (в т. ч. предшествующее лечение антикоагулянтами);
• воспалительные заболевания женских половых органов;
• наличие тяжелой экстрагенитальной патологии.
• наличие в анамнезе повышенной чувствительности к мифепристону;
• надпочечниковая недостаточность и длительная глюкокортикостероидная терапия;
• острая или хроническая почечная и/или печеночная недостаточность;
• порфирия;
• анемия;
• нарушения гемостаза (в т. ч. предшествующее лечение антикоагулянтами);
• воспалительные заболевания женских половых органов;
• наличие тяжелой экстрагенитальной патологии.
В результате проведенных исследований было показано, что при ежедневном применении мифепристона в дозе 50 мг у 30 пациенток размер ЛММ (по данным ультразвукового исследования – УЗИ) уменьшался в следующем процентном соотношении:
• через 4 недели после начала исследования – на 20,8–44,1 % от размера ЛММ до начала лечения;
• через 8 недель – на 29,8–51,2 % от размера ЛММ до начала лечения;
• через 12 недель – на 47,3–58,3 % от размера ЛММ до начала лечения.
• через 4 недели после начала исследования – на 20,8–44,1 % от размера ЛММ до начала лечения;
• через 8 недель – на 29,8–51,2 % от размера ЛММ до начала лечения;
• через 12 недель – на 47,3–58,3 % от размера ЛММ до начала лечения.
Подтверждением эффективности применения препарата мифепристон явилось также уменьшение в среднем на 10,2 % интенсивности кровообращения в миоматозных узлах через 3 месяца после начала лечения, по данным допплерометрии, по сравнению с первоначальными значениями. Ежедневное применение мифепристона в дозе 25 мг у 30 пациенток с множественной ЛММ показало, что размер ЛММ, по данным УЗИ, уменьшался в следующем процентном соотношении:
• через 4 недели от начала исследования – на 17,9–42,3 % от размера ЛММ до начала лечения;
• через 8 недель – на 27,9–49,8 % от размера ЛММ до начала лечения;
• через 12 недель – на 43,7–56,4 % от размера ЛММ до начала лечения.
• через 4 недели от начала исследования – на 17,9–42,3 % от размера ЛММ до начала лечения;
• через 8 недель – на 27,9–49,8 % от размера ЛММ до начала лечения;
• через 12 недель – на 43,7–56,4 % от размера ЛММ до начала лечения.
Интенсивность кровообращения,по данным допплерометрии, через 3 месяца после лечения уменьшилась в среднем на 12,4 % по сравнению с первоначальными значениями.
У 92 % от общего числа пациенток во время лечения наблюдалась аменорея, в 8 % случаев – олигоменорея, вызванные применением мифепристона. Другие побочные эффекты препарата отсутствовали.
Как показало исследование крови, уровень таких гормонов, как ЛГ, эстрадиол, изменялся после лечения, но статистически недостоверно по сравнению с контролем. Уровень прогестерона после лечения по сравнению с контролем уменьшился. В клиническом анализе крови, проводимом
как во время лечения, так и после его окончания, отклонений от нормы не было. Аллергической реакции на мифепристон или его индивидуальной непереносимости ни у одной из пациенток выявлено не было.
как во время лечения, так и после его окончания, отклонений от нормы не было. Аллергической реакции на мифепристон или его индивидуальной непереносимости ни у одной из пациенток выявлено не было.
Авторы сделали следующие выводы:
1. Применение мифепристона в монотерапии ЛММ, соответствующей размерам до 12 недель беременности, является эффективным и безопасным методом лечения.
2. В период исследования и дальнейшего наблюдения (в течение 6 месяцев) осложнений лечения не отмечено.
3. Дозы мифепристона 50 и 25 мг перорально ежедневно в течение 3 месяцев являются эффективными.
4. Мифепристон может быть рекомендован в качестве средства для терапевтического лечения ЛММ до 12 недель беременности.
1. Применение мифепристона в монотерапии ЛММ, соответствующей размерам до 12 недель беременности, является эффективным и безопасным методом лечения.
2. В период исследования и дальнейшего наблюдения (в течение 6 месяцев) осложнений лечения не отмечено.
3. Дозы мифепристона 50 и 25 мг перорально ежедневно в течение 3 месяцев являются эффективными.
4. Мифепристон может быть рекомендован в качестве средства для терапевтического лечения ЛММ до 12 недель беременности.
Таким образом, терапия ЛММ мифепристоном по эффективности и безопасности не уступает аналогам аГРГ, при этом не обладает отрицательным воздействием на эстрогензависимые экстрагенитальные системы и органы (сердечно-сосудистую систему, костную ткань, молочные железы и т. д.), поскольку сывороточный уровень эстрадиола остается на уровне, соответствующем ранней фолликулярной фазе.
Интерес к мифепристону не ослабевает, и в настоящее время в США проходит IIА стадия клинических испытаний мифепристона (таблетки 10 мг вагинально однократно в течение 3 месяцев) в лечении ЛММ (NCT00881140), результаты которого будут опубликованы в 2011 г.
Кроме мифепристона в качестве антигестагенных препаратов в терапии ЛММ используются азоприснил и улипристел, пока не зарегистрированные в РФ. В частности, закончилась IIВ фаза клинических испытаний селективного модулятора РП улипристела ацетата (Ulipristal acetate, CDB-2914,
PGL4001) в лечении миомы матки (www.clinicaltrials.gov). Двойное слепое плацебо-контролируемое исследование продемонстрировало высокую эффективность улипристела в сокращении объема матки и миомы с достоверным повышением качества жизни пациенток (40 женщин, 12 недель). В настоящее время проходит III стадия испытаний (PEARL I&II, 14 стран, 540 женщин), результаты которого будут представлены в середине 2010 г. Однако в РФ эти препараты пока не зарегистрированы.
PGL4001) в лечении миомы матки (www.clinicaltrials.gov). Двойное слепое плацебо-контролируемое исследование продемонстрировало высокую эффективность улипристела в сокращении объема матки и миомы с достоверным повышением качества жизни пациенток (40 женщин, 12 недель). В настоящее время проходит III стадия испытаний (PEARL I&II, 14 стран, 540 женщин), результаты которого будут представлены в середине 2010 г. Однако в РФ эти препараты пока не зарегистрированы.
Клинические испытания проведены для мифепристона в дозах 25 и 50 мг [6]. Зарегистрирорван в РФ и применяется в лечебной практике с 2005 г. препарат Гинестрил (мифепристон 50 мг для перорального приема). Гинестрил назначают по 1 табл./сут в течение 3 месяцев.
Заключение
Таким образом, клиническая эффективность и хорошая переносимость мифепристона позволяют успешно использовать его при лечении ЛММ у женщин репродуктивного возрата [11]. Дальнейшие исследования, посвященные изучению клинической эффективности мифепристона при вариации его доз и путей введения в зависимости от состояния рецепторного аппарата миометрия и миоматозных узлов, необходимы для персонализованного применения данного препарата в лечении ЛММ.



{kind=link}
{kind=link}
{kind=link}
{kind=link}